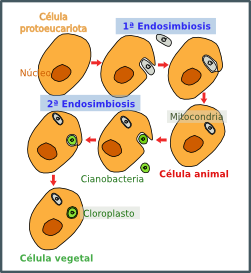
kali ini kita akan bahas sejarah teori endosimbiosis,dari prokariot ke eukariot, dari aerob ke anaerob dan dari tidak berkloroplas dan mitokondria, sampai akhirnya terbentuk sel dengan kloroplas dan mitokondria ......
Transisi dan
evolusi antara eukariot,sel dengan inti dan prokariot, menurut beberapa ahli
menjadi perubahan paling mendalam dalam
sejarah evolusi,asal-usul dari sel eukariot yang menurut beberapa ahli berasal
dari aktvitas endosymbiosis, yang mena dalam teori ini menjelaskan fenomena
adanya organisme yang hidup dalam satu
komponen organisme saling melengkapi satu sama lain beberapa orang lebih
mengenal dengan istilah hipotesis endosymbiosis, hipotesis ini menyebutkan
bahwa sel eukariotuk berasal dari perkembangan sel prokariotik
Hal ini didasarkan
pada kenyataan struktur dari sel eukariotik :
-
Merupakan
membrane rangkap,terdiri dari membrane luar dan membrane dalam
-
Membrane
luar nucleus memiliki hubungan secara langsung dengan membrane sel reticulum
endoplasma.Hubungan ini merupakan sisa-sisamembran plasma yang melekuk ke dalam
Gagasan bahwa pada dasarnya sel eukariotik sebenarnya
merupakan koloni mikroba pertama kali diusulkan pada tahun 1920 oleh ahli
biologi Amerika Ivan Wallin (Fausto-Sterling 1993). Pencetus versi modern SET
adalah biologi Lynn Margulis. Pada tahun 1981,Margulius dengan edisi pertama
yang berjudul simbiosis eolution, dimana awala gagasanya menyatakan bahwa sel
eukariotik berinterkasi,bergabung dalam urutan tertentu. Nenek moyang dahulunya
mendapatkan organel tersebut dari dalam sel inangnya baik yang menjadi
mangsanya maupun parasite internal,selanjutnya hubungan itu berkembang menjadi
hubungan saling menguntungkan bagi kedua organisme,saling bergantung dan
akhirnya berevolusi.
Dalil bahwa nenek moyang mitokondria yang
hidup bebas , seperti halnya Daptobacter dan Bdellovibrio, yang mengembangkan
kemampuan untuk secara efisien bernafasdengan memanfaatkan oksigen (Margulis dan Sagan 1987).
Itu dapat dibuktikan dengan adanya struktur
dari mitokondria :
-
Mitokondria
memilik dua membrane yaitu membrane luar dan membrane dalam.membran luar diduga
membrane sel inang yang melekuk kedalam, ketika menelan sel bakteri
aerob,sedangkan membrane dalam diduga merupakn membrane bakteri aerob
-
Bukti
selanjutnya yiatu bahkan sampai saat ini ada bakteri aerob yang memilikimesosom
sebagai penghasil energy, diduga sel prokariot anaerob mirip dengan bakteri
aerob tersebut
-
Didalam
mitokondria terdapat DNA yang mengontrol pembuatan polipeptida yang berbeda dengan
DNA Inang, diduga mirip dengan DNA prokariot
-
Polipeptida
yang dihasilkan didalam mitokondria digunakan sendiri oleh mitokondria
tersebut, polipeptida ini berbeda dengan polipeptida sel inang
-
Mitokondria
mampu membelah diri seperti bakteri sehingga jumlahnya dapat bertambah banyak
Sedangakan
nenek moyang kloroplas hari ini adalah cyanobacteria, yang awalnya organisme
fotosintetik independen. Selain itu, silia whiplike yang umum pada eukariota
tetapi tidak ditemukan pada prokariota yang diperkirakan berasal dari kelompok
lain masih hidup bebas . Menurut SET, sel host asli prokariotik adalah
archaebacterium, mirip dengan Thermoplasma yang ditemukan hari ini, yang dapat
menahan suhu tinggi dan kondisi asam (Margulis dan Sagan 1987). Ini adalah sel
inang tidak berfotosintesis dan mampu
secara efektif menggunakan oksigen.
Hipotesis untuk pemebntukan kloroplas ini
sendiri dpada dasarnya dikemukakan berdasarkan bahwa :
-
Kloroplas
meiliki membrane ganda dan membrane luarnya mrip dengan struktur membrane sel
-
Ada
bakteri fotosintetik yang memliki membrane fotosintetik mirip dengan tilakoid
pada kloroplas
-
Didalam
kloroplas terdapat DNA yang juga dijumpai pada bakteri fotosintetik.DNA
kloroplas memiliki fungsi versi tersendiri tidak dipengaruhi dNA nucleus sel inang.
-
Kloroplas
dapat bertambah banyak melalui pembelahan seperti halnya bakteri
Sepanjang tulisannya, Margulis menyatakan simbiosis yang merupakan
kekuatan pendorong utama di balik evolusi. Menurutnya, kerja sama, interaksi,
dan saling ketergantungan di antara bentuk kehidupan diperbolehkan untuk
dominasi global yang akhirnya hidup. Akibatnya, gagasan evolusi Darwin sebagai
"survival of the fittest," adalah sebuah kompetisi yang terus-menerus
antara individu dan spesies, dinilai tidak lengkap. Menurut Margulis dan Sagan
(1986), "Hidup tidak mengambil alih dunia dengan pertempuran, tetapi oleh
jaringan." Daripada fokus semata-mata pada penghapusan pesaing, pandangan
Margulis lebih mengutamakan simbiosis
Penemuan awal yang penting dalam mendukung SET terjadi di laboratorium
Kwang W. Jeon, seorang ahli biologi University of Tennessee. Jeon menyaksikan
pembentukan simbiosis amuba-bakteri simbion bakteri yang baru menjadi
terintegrasi dalam host amuba (Jeon 1991). Pada tahun 1966, saat pertama kali
terinfeksi bakteri amuba, mereka mengaplikasikannya untuk host ataupun bahan
percobaan mereka. Namun, seiring berjalannya waktu, beberapa amuba terinfeksi
selamat dan menjadi tergantung pada endosymbion. Jeon membuktikan
integrasi ini dengan melakukan
transplantasi inti antara amuba terinfeksi dan amuba netral . Jika dibiarkan
saja, para amuba hybrid meninggal dalam hitungan hari. Namun jika ia merupakan
bentukan hibrida ini dengan simbiosis, amuba pulih dan hidup serta kembali
berkembang (Margulis dan Sagan 1987). Penemuan ini berfungsi untuk menunjukkan
mekanisme utama pembentukan el baru yang terbentuk melauli system
endosimbiosis(Jeon 1991).
Meskipun beberapa ide Margulius 'tetap kontroversial, ada banyak bukti
yang mendukungnya. Bukti ini berfungsi untuk mempertahankan gagasan asal
endosymbiotic mitokondria dan kloroplas. Pengakuan bahwa kloroplas mitokondria
dan bisa muncul hanya dari yang sudah ada sebelumnya. Para ilmuwan menemukan
bahwa mitokondria dan kloroplas tidak dapat dibentuk dalam sel yang tidak memiliki
struktur mereka karena kode dan struktur yang berbeda . Juga, baik mitokondria
dan kloroplas memiliki set mereka sendiri ,gen yang lebih mirip dengan
prokariota daripada eukariota. Mereka berdua mengandung molekul DNA melingkar,
seperti yang ditemukan di prokariota. Akhirnya, baik mitokondria dan kloroplas
memiliki sendiri protein-sintesis mesin.
Aspek paling
diterima dan paling dipertanyakan dari SET adalah hipotesis undulipodia
eukariotik berasal dari bakteri spirochete (Margulis 1993). Istilah "undulipodia"
digunakan untuk menggambarkan motilitas organel eukariotik, flagela dan silia.
Undulipodia terdiri dari mikrotubulus dalam konfigurasi tertentu. Mikrotubulus
yang terdiri dari protein yang terkait erat beberapa disebut Tubulin. Struktur ini
jauh lebih besar dan lebih kompleks daripada flagela bakteri, yang terbuat dari
protein flagellin. SET ini mendalilkan bahwa undulipodia mungkin berasal dari
bakteri melalui simbiosis motilitas (Bermudes, Margulis, dan Tzertzinis 1987).
Ide ini disebut sebagai hipotesis eksogen. Rincian argumen yang kompleks,
tetapi pendukung titik SET untuk beberapa baris bukti. Argumen mereka
menekankan biologi organel sendiri, distribusi mereka, dan terjadinya struktur
terkait dan analog. Para penentang pandangan ini, pendukung hipotesis endogen,
menunjukkan bahwa undulipodia berasal internal sebagai perpanjangan
mikrotubulus digunakan dalam mitosis. Hipotesis ini juga disebut sebagai
filiasi langsung, yang merupakan pandangan nonsimbiotik evolusi yang menekankan
peran berbagai jenis mutasi pada pemisahan evolusi sel eukariotik dari sel
prokariotik.
Kontroversi utama antara hipotesis endogen dan eksogen untuk asal undulipodia
bersandar pada soal kronologi. Para pendukung klaim hipotesis endogen mengklaim
mikrotubulus mendahului asal undulipodia, yang akhirnya muncul bentukan
endogen. Sebaliknya, negara-negara hipotesis eksogen yang simbiosis motilitas
memunculkan sel dengan undulipodia, dan akuisisi ini kemudian menyebabkan
struktur internal yang terlibat dalam mitosis (Bermudes, Margulis, dan
Tzertzinis1987). Meskipun asal simbiosis undulipodia adalah mendapatkan
dukungan, kontroversi masih harus diselesaikan. Menurut Bermudes dan Margulis
(1985), terdapat bukti yang cukup untuk membuktikan baik filiasi langsung atau
hipotesis simbiosis untuk asal undulipodia.
Untuk memahami implikasi teori
klasifikasi dari semua bentuk kehidupan, ringkasan singkat dari
penafsiran saat peristiwa endosymbiotic diperlukan. Menurut teori ini,
eukariota berkembang ketika sel-sel archaea dan eubacterial bergabung dalam
simbiosis anaerobik. Sel archaea disediakan sitoplasma sel sedangkan
eubacterial (spirochete a) diperbolehkan untuk mobilitas dan, pada akhirnya,
mitosis…
Meskipun inti adalah karakteristik yang mendefinisikan sel eukariotik, asal
usul organel ini dan simbiosisnya tidak
pasti,. Margulis cenderung mendukung proses yang melibatkan kombinasi filiasi
langsung dan simbiosis sebagai sumber dari sel berinti. Dia percaya bahwa
beberapa sel prokariotik berevolusi inti primitif melalui filiasi langsung
tetapi tetap prokariotik. Lainnya berevolusi struktur yang sama tetapi juga
diperoleh gen simbiosis dan akibatnya
menjadi eukariota (Margulis 1993).
Sebuah makalah tahun 1996 oleh Golding dan Gupta , sengketa pandangan
tradisional tentang asal-usul inti dan menyarankan alternatif yang disebut
model chimeric. Istilah "chimeric" merujuk pada suatu organisme yang
mengandung jaringan dari setidaknya dua orang tua yang berbeda secara genetik.
Model chimeric mengusulkan bahwa sel eukariotik pertama muncul sebagai hasil
dari peristiwa fusi yang tidak biasa antara eubacterium Gram-negatif (host)
tanpa dinding sel dan archaebacterium (Simbion) di mana kedua orang tua membuat
kontribusi besar untuk genom nuklir sel. Inti muncul sebagai hasil lipat di
membran inang sekitar sel ditelan. Model chimeric didasarkan pada bukti genetik
dan biokimia. Salah satu bukti genetik yang mendukung model adalah fakta bahwa
sel-sel prokariotik yang homogenomic (memiliki materi genetik dari salah satu
orang tua saja), sedangkan sel eukariotik yang heterogenomic (memiliki materi
genetik dari lebih dari satu orang tua). Bukti biokimia dalam mendukung model
chimeric melibatkan filogenetik, atau evolusi, analisis data sekuens dari
protein. Analisis ini menunjukkan hubungan erat antara bakteri Gram-negatif dan
eukariota di satu sisi dan Gram-positif bakteri dan archaebacteria di sisi lain
(Golding dan Gupta 1996). Data protein bahkan lebih urutan menyarankan hubungan
antara eukariota dan archaebacteria. Data ini menunjukkan bahwa hubungan
simbiosis antara bakteri Gram-negatif dan archaebacteria sebagai nenek moyang
dari sel eukariotik layak. Secara keseluruhan, data sekuens mendukung model
chimeric.
Penelitian Martin dan Müller
(1998) ke asal mitokondria telah menyebabkan teori baru yang disebut
endosimbiosis Pada gambar saat asal dari sel eukariotik "hipotesis
hidrogen.", Mitokondria adalah "kecelakaan beruntung" (Vogel
1998). Sel inang leluhur hanya menelan nenek moyang mitokondria, tidak
sepenuhnya menelan itu, dan sebuah sel bahkan lebih sukses menghasilkan.
Menurut hipotesis hidrogen, bagaimanapun, sel eukariotik pertama tidak
terbentuk hanya secara kebetulan. Sebaliknya, itu adalah hasil dari kesatuan
tujuan antara sel inang archaebacterial, sinkronisasi yang akhirnya menghasilkan metana, dan simbion mitokondria masa depan
yang membuat hidrogen dan karbon dioksida sebagai produk limbah dari
metabolisme anaerobik. Dengan demikian, meskipun Simbion itu mungkin mampube
respirasi aerobik, simbiosis dimulai dari hasil dari produk metabolisme
anaerobik. Ketergantungan tuan rumah pada hidrogen diproduksi oleh simbion
tersebut diidentifikasi sebagai prinsip selektif yang mengkonsolidasi nenek
moyang sel eukariotik (Martin Müller dan 1998).
Hipotesis hidrogen memiliki beberapa implikasi penting yang bertentangan
dengan pandangan saat ini hubungan antara eukariota dan archaebacteria. Dalam
pandangan saat ini, eukariota bercabang dari sejak lama dari archaebacteria
sebelum archaebacteria telah dibagi menjadi kelompok. Hipotesis hidrogen menyiratkan bahwa
eukariota pertama muncul,kemudian pada gambar mekanismeevolusi, yang
menggambarkan kaitan erat terhadap archabacteria.
Penjelasan lain baru-baru ini tentang asal-usul eukariota yang disebut
"hipotesis syntrophic" disajikan oleh García López dan Moreira
(1998). Meskipun mereka secara independen mengusulkan, hipotesis syntrophic
bersifat komplementer dalam beberapa aspek dengan hipotesis hidrogen. Kedua
hipotesis setuju saran dari metabolisme anaerobik untuk asal simbiosis mitokondria.
Mereka juga sangat mirip dalam beberapa rincian metabolisme simbiosis dan fitur
molekul archaea (López-Garcia dan Moreira 1998). Perbedaan utama antara dua
hipotesis adalah sifat kemitraan bakteri asli. Seperti yang dinyatakan
sebelumnya, dalam hipotesis hidrogen, simbiosis asli diperkirakan terjadi
antara archaebacterium metanogen dan nenek moyang eubacterial ke mitokondria.
Dalam hipotesis syntrophic, simbiosis aslinya dipahami telah terjadi antara
archaebacterium metanogen dan sulfat-respiring leluhur delta-proteobacterium.
Yang pertama memberikan materi genetik pusat dan metabolisme asam nukleat
sedangkan yang kedua diberikan karakteristik yang paling metabolik
(López-Garcia dan Moreira 1998). Mitokondria diperkirakan berasal dari acara,
kemudian simbiosis independen. Seperti dengan hipotesis hidrogen, analisis
sekuensing genetik lanjut diperlukan dalam rangka untuk klaim hipotesis
syntrophic harus ditegakkan.
Hampir tiga puluh tahun sejak Lynn Margulis pertama kali diterbitkan sebuah
buku tentang asal-usul sel eukariotik. Sejak saat itu, biologi telah mengalami
perubahan yang luar biasa. Perubahan yang paling terlihat adalah akumulasi
ekstensif data sekuens untuk kedua asam nukleat dan protein. Pengumpulan data
baru niscaya akan mengakibatkan revisi terus menerus dari teori endosimbiosis
serial asal dari sel eukariotik. Meskipun ketidakpastian masa depan, fondasi
penting telah diletakkan. Simbiosis kini diterima oleh komunitas ilmiah sebagai
faktor penting dalam menghasilkan perubahan evolusioner. Langkah berikutnya
termasuk pengembangan metode yang lebih rumit untuk menafsirkan data sekuens
genetik dan molekuler dan usaha dari segar melihat catatan fosil.
Posted By : Muhammad Rezki Rasyak